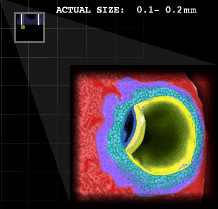
[Como se ha anunciado en el artículo precedente, publicamos ahora la segunda parte de un completo texto sobre las células madre y los embriones humanos preparado por las Profesoras Natalia López Moratalla e (cfr. palabras introductorias a # 196). La figura en espiral que se incluye sobre estas líneas - The Visible Embryo - representa primero (en azul) las 23 etapas que ocurren en el primer trimestre del embarazo; y luego (en verde y en morado), el continuo crecimiento del embrión humano concretado cada dos semanas a lo largo del segundo y tercer trimestre, hasta llegar el momento del nacimiento. Es un proceso continuo que se inicia en el momento de la fecundación. Como todo científico sabe, ya a los "pocos días" esa “masa celular” concreta, no es sin más un conjunto de células sino un "embrión humano completo de pocos días". Por tanto, no hay que estar especialmente dotado intelectualmente para llegar a la conclusión de que si se toman las células de los "embriones sobrantes de la fecundación in vitro", o se producen embriones para usarlos como medio de investigación, significa destruir vidas humanas. Como en aquel antiguo chiste, también en esto hay que hablar muy claro y "no llamar al pan, zuchf, y al vino, frolo..."]
[Como se ha anunciado en el artículo precedente, publicamos ahora la segunda parte de un completo texto sobre las células madre y los embriones humanos preparado por las Profesoras Natalia López Moratalla e (cfr. palabras introductorias a # 196). La figura en espiral que se incluye sobre estas líneas - The Visible Embryo - representa primero (en azul) las 23 etapas que ocurren en el primer trimestre del embarazo; y luego (en verde y en morado), el continuo crecimiento del embrión humano concretado cada dos semanas a lo largo del segundo y tercer trimestre, hasta llegar el momento del nacimiento. Es un proceso continuo que se inicia en el momento de la fecundación. Como todo científico sabe, ya a los "pocos días" esa “masa celular” concreta, no es sin más un conjunto de células sino un "embrión humano completo de pocos días". Por tanto, no hay que estar especialmente dotado intelectualmente para llegar a la conclusión de que si se toman las células de los "embriones sobrantes de la fecundación in vitro", o se producen embriones para usarlos como medio de investigación, significa destruir vidas humanas. Como en aquel antiguo chiste, también en esto hay que hablar muy claro y "no llamar al pan, zuchf, y al vino, frolo..."] ___________________________
IV.- Células pluripotentes embrionarias
El contexto histórico
Diversos trabajos se han dirigido hacia la diferenciación de células madre embrionarias pluripotentes humanas, aisladas de la masa celular interna del embrión en los primeros días de desarrollo (blastocisto), que se han diferenciado in vitro hacia una amplia variedad de tipos celulares (Thomson, J.A., et al., 1998; Shamblott M.J., et al. 1998; Nagy A., et al., 1993). Estas células muestran, en condiciones de cultivo adecuadas, una ilimitada capacidad de multiplicarse y al mismo tiempo que mantienen la capacidad de dar diferenciación original. En 1998, James Thomson y su equipo describieron un método para el mantenimiento in vitro de células madre embrionarias derivadas de la masa celular interna de blastocistos humanos producidos mediante fecundación in vitro, donados para investigación por las clínicas de reproducción asistida. Al mismo tiempo, otro grupo, dirigido por John Gearhart, publicó experimentos similares, con la derivación de células embrionarias germinales, obtenidas de gónadas y tejido mesenquimático de fetos abortados de 5 a 9 semanas.
Ambos grupos desarrollaron su investigación gracias a experimentos previos que se remontan al siglo XIX; en efecto ya en 1878, se publica el primer intento de fertilización de óvulos de mamíferos fuera del organismo y en 1959, el primer artículo de ratones producidos mediante fecundación in vitro en los Estados Unidos. En 1968, se consigue la primera fecundación de óvulos humanos en el laboratorio y, en 1978, nace en el Reino Unido, Louise Brown, el primer bebé producido mediante fecundación in vitro. En los años 80 se establecen las primeras condiciones de cultivo para células madre embrionarias derivadas de la masa celular interna de blastocistos de ratones. Y se desarrollan, a partir de una línea celular de teratocarcinoma testicular, células pluripotentes, denominadas carcinoma embrionario, que expuestas a ácido retinoico eran capaces de diferenciarse a células similares a neuronas y otros tipos celulares.
En 1994 se generan blastocistos humanos, con fines reproductivos, en el laboratorio y comienzan los trabajos encaminados a mantener, en cultivo, las células de la masa interna. En los años siguientes se obtienen células madres de primates, a partir la masa celular interna de blastocistos de mono rhesus.
Las células aisladas de la masa celular interna de blastocistos humanos se cultivaron en un estrato de fibroblastos de ratón irradiados, donde se multiplican y confluyen hasta la formación de colonias llamadas "cuerpos embrioides" (Itskovitz-Eldor, J., 2000) con las tres capas germinales. Por cultivos repetidos de las células de las colonias obtenidas se consigue una línea con capacidad de multiplicación indefinida que conserva las características de células troncales durante meses y años.
Características de las células embrionarias
Las células humanas provenientes de la masa interna de blastocistos, aisladas y cultivadas en el año 98, por Thomson y su equipo, presentan cariotipos normales, se mantienen viables a lo largo de múltiples multiplicaciones. Mantienen la telomerasa (Betts D.H. y King W.A. 1999) por lo que se replican de forma indefinida y se diferencian reguladas por los factores del medio (Watt F.M. y Hogan B.L., 2000).
Se han conseguido líneas celulares a partir de las del embrión temprano (H9.1 y H9.2) que retienen las propiedades de estas incluida la capacidad de proliferar, generar tumores teratomas in vivo, y diferenciarse a células que derivan de las tres capas del embrión de las que se desarrollan todos los tipos celulares (Amit M., et al., 2000).
Más aún, las células madre del embrión expresan en la membrana antígenos característicos de los primeros estados del desarrollo embrionario, tales como el SSEA-3 y SSEA-4 (Thomson J.A. y Marshall V.S., 1998), que mantienen las interacciones especificas entre ellas, por lo que cuando se retiran de los lechos biológicos en que crecen y se ponen en cultivo forman, entre los 7 y 14 días, embriones en estado de mórula y blastocisto y diferencian un trofoblasto (Itskovitz-Eldor J., et al. 2000).
Estas características muestran lo que es obvio: esta “masa celular” concreta, la del embrión de unos cinco días, no es sin más un conjunto de células sino un embrión completo de pocos días. Tomar sus células de los embriones “sobrantes de la fecundación in vitro” o producirlos directamente para usarlos, es destruirlos.
Esas células de la masa interna, incluso después de hacerlas multiplicarse en el laboratorio son capaces de formar la capa celular que las recubre, el trofoblasto. Por ello, querríamos llamar la atención hacia este hecho; en las circunstancias de cultivo que se han mostrado en la primera figura, lo que han llamado cuerpos embrioides, son realmente embriones “clónicos” del embrión producido por fecundación in vitro, y usado como punto de partida. En las condiciones de laboratorio estos cuerpos embrioides no tienen ninguna posibilidad de desarrollarse y de hecho no se sabe, porque no se les ha dado la oportunidad de anidar en el útero de una mujer, si son verdaderos embriones tempranos capaces de dar un organismo completo. No haber comprobado esta posibilidad con animales es otra muestra de que el interés por las cotizadas células madre embrionarias humanas, pasa por encima de cualquier reserva ética hacia la vida humana en sus inicios.
Potencial terapéutico de las células madre embrionarias
La potencialidad de las células madre del embrión parece muy amplia. Las células madre del embrión se diferencian para dar una gran variedad de tipos celulares: cardiomiocitos, progenitores hematopoyéticos, miocitos esqueléticos (Doetschman T.C., et al., 1985), células musculares (Baker R.K., y Lyons G.E.,1996), adipocitos (Dani C., 1999), condriocitos (Poliard A., et al., 1995) células endoteliales (Risau W., et al., 1988), melanocitos (Yamane T., et al., 1999), neuronas y células de la glía (Brustle O., et al., 1999; Bain G., et al., 1995) y por último células de los islotes beta pancreáticos (Soria B., et al., 2000).
Ahora bien, insistimos, el uso terapéutico de células madre procedentes de embriones de 5 días, en estado de blastocisto, supondrá siempre la destrucción de estos embriones humanos. Por otra parte, las informaciones divulgadas han llevado a concluir erróneamente que los científicos estaban a punto de dominar la síntesis artificial de órganos para trasplantes, y con ello el debate suscitado por la destrucción de embriones continúa presentando, y urgiendo, esta investigación como el mejor método para curar enfermedades que como la de Parkinson, Alzehimer, la diabetes o el infarto de miocardio padecen un elevado número de personas.
Sin embargo, y además de los inconvenientes éticos, justamente por la pluripotencialidad de las células madre embrionarias, están sin resolver las técnicas que permitan dirigirlas en la dirección deseada para el transplante terapéutico y también está por controlar su proliferación indeterminada (Shamblott M.J., et al., 2001); de hecho, provocaron tumores en los experimentos realizados en animales.
Más aún estas células provocan rechazo inmunológico por parte del receptor, por provenir de un donante diferente genéticamente. Para eliminar el rechazo no es del todo adecuado usar las terapias inmunosupresoras por el peligro de no poder hacer frente a infecciones o combatir posibles tumores en una situación de bajas defensas del organismo. Se trabajan diferentes vías (Kaufman D.S., et al., 2000) como eliminar los antígenos MHC extraños, o reemplazarlos (Grusby M.J. et al., 1993), o usar, como veremos después, la clonación por transplante del núcleo de una células del paciente para generar una línea de células madre del embrión clonado compatible inmunológicamente con el paciente de cuya célula se tomó el material genético nuclear (First N.L. y Thomson J., 1998).
Aunque en el mes de julio de 2001 un grupo de investigadores estadounidenses, del Instituto Jones en el estado de Virginia, ha creado embriones humanos y los ha destruido después de unos días para aislar las células madre pluripotentes, lo habitual hasta entonces había sido usar para este fin los embriones “sobrantes” de la fecundación in vitro. Veremos más adelante porqué los embriones humanos procedentes de la fecundación de un óvulo obtenido en un proceso de estimulación ovárica encaminada a una multiovulación resultan alterados: menor capacidad de implantación en el útero y malformaciones en diversos órganos.
_________________________
Bibliografía (IV)
Amit, M., Carpenter, M.K., Inokuma, M.S. et al. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev. Biol. 2000; 227:271-278.
Baker, R.K., Lyons, G.E. Embryonic stem cells and in vitro muscle development. Curr. Top Dev. Biol. 1996; 33:263-279.
Bain, G., Kitchens, D., Yao, M. et al. Embryonic stem cells express neuronal properties in vitro. Dev. Biol. 1995; 168:342-357.
Betts, D.H., King, W.A. Telomerase activity and telomere detection during early bovine development. Dev. Genet. 1999; 25:397-403.
Brustle, O., Jones, K.N., Learish, R.D. et al. Embryonic stem cell-derived glial precursors: a source of myelinating transplants. Science 1999; 285:754-756.
Dani, C. Embryonic stem cell-derived adipogenesis. Cells Tissues Organs 1999; 165:173-180.
Doetschman, T.C., Eistetter, H., Katz, M. et al. The in vitro development of blastocyst-derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J. Embryol. Exp. Morphol. 1985; 87:27-45.
Grusby, M.J., Auchincloss, H. Jr., Lee, R. et al. Mice lacking major histocompatibility complex class I and class II molecules. Proc. Natl. Acad. Sci. USA 1993; 90:3913-3917.
First, N.L., Thomson, J. From cows stem therapies? Nat. Biotechnol. 1998; 16:620-621.
Itskovitz-Eldor, J., Schulding, M., Karsenti, D. et al. Differentiation of human embryonic stem cells into embryoid bodies comprising the three embryonic germ layers. Mol. Med. 2000; 6:88-95.
Kaufman, D.S., Odorico J.S., Thomson, J.C. Transplantation therapies from human embryonic stem cells circumventing immune rejection. e-biomed 2000;1:11-15: available at http://www.liber/pub.com/EBI/defaultstatic.asp).
Nagy, A., Rossant, J., Nagy, R. et al. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proc. Natl. Acad. Sci. USA 1993; 90:8424-8428.
Poliard, A., Nifuji, A., Lamblin, D. et al. Controlled conversion of an immortalized mesodermal progenitor cell towards osteogenic, chondrogenic, or adipogenic pathways. J. Cell Biol. 1995; 130:1461-1472.
Risau, W., Sariola, H., Zerwes, H.G. et al. Vasculogenesis and angiogenesis in embryonic-stem-cell-derived embryoid bodies. Development 1988; 102:471-478.
Shamblott, M.J., Axelman, J., Littlefield, J.W. et al. Human embryonic germ cell derivatives express a broad range of developmentally distinct markers and proliferate extensively in vitro. Proc. Natl. Acad. Sci. USA 2001; 98:113-118.
Shamblott, M.J., Axelman, J., Wang, S. et al. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc. Natl. Acad. Sci. USA 1998; 95:13726-13731.
Soria, B., Roche, E., Berna, G. et al. Insulin-secreting cells derived from embryonic stem cells normalize glycemia in streptozotocin-induced diabetic mice. Diabetes 2000; 49:157-162.
Thomson, J.A., Itskovitz-Eldor, J., Shapiro, .S.S. et al. Embryonic stem cell lines derived from human blastocysts. Science 1998; 282:1145-1147.
Thomson, J.A., Marshall, V.S. Primate embryonic stem cells. Curr. Top Dev. Biol. 1998; 38:133-165.
Watt, F.M., Hogan, B.L. Out of Eden: Stem cells and their niches. Science 2000; 287:1427-1430.
Wilmut, I., Schnieke, A.E., McWhir, J. et al. Viable offspring derived from fetal and adult mammalian cells. Nature 1997; 385:810-813.
Wobus, A.M., Wallukat, G., Hescheler, J. Pluripotent mouse embryonic stem cells are able to differentiate into cardiomycocytes expressing chronotropic responses to adrenergic and cholinergic agents and Ca2+ channel blockers. Differentiation 1991; 48:173-182.
Yamane, T., Hayashi, S., Mizoguchi, M. et al. Derivation of melanocytes from embryonic stem cells in culture. Dev. Dyn. 1999; 216:450-458.
____________________________________
V.- Células madre germinales fetales.- Clonación y partenogénesis de óvulos.
Células madre pluripotentes de fetos humanos
En los experimentos del equipo de Gearhart (Shamblott et al., 1998) las células madre germinales se aislaron de fetos abortados; concretamente de la región del feto destinada a desarrollarse a gónadas y formar los gametos. A pesar de la diferente fuente de obtención, las células madre pluripotenciales que se obtuvieron son muy similares a las aisladas del blastocisto. Las células madre germinales, extraídas de fetos, son también pluripotenciales, capaces de dar en cultivo en el laboratorio diversos tipos de células maduras.
Estas células fetales se han usado ya en un intentos de paliar la enfermedad de Parkinson. Los ensayos con ratas en este sentido han demostrado que si se insertan células pluripotenciales, tratadas previamente para que produzcan niveles altos de dopamina, se consigue una reducción en los síntomas de dicha lesión neurológica (Studer, L.et al. 1998). No obstante, la necesidad de aumentar el control de esos injertos, para su uso en humanos es patente. López Lozano, en Madrid, ha efectuado desde 1988, trasplantes de tejido de mesencéfalo de fetos a pacientes con Parkinson (López-Lozano, J.J., et al., 2000); de los 42 pacientes a los que se ha realizado este tipo de intervención, el 60% han mostrado una mejoría clínica en un período de más de 7 años. Pero, en el primer estudio a doble ciego realizado en 40 pacientes de Parkinson, (Freed C.R., et al. 2001) se ha comprobado que es muy escaso el efecto beneficioso que las células fetales inyectadas en el cerebro de pacientes con Parkinson tienen sobre la evolución clínica de su enfermedad: consiguieron en los pacientes más jóvenes, menores de 60 años, un incremento del 20% en la producción de dopamina y una reducción en los síntomas clínicos del Parkinson, que se mantenía a los 36 meses del trasplante; pero no se obtuvo mejoría en los pacientes de edad más avanzada, incluso en éstos los efectos fueron negativos (en un 15% de los pacientes se dio una superproducción de células fetales, y fabricaron a su vez tal cantidad de una sustancia química relacionada con el movimiento, que los pacientes se retorcían y se sacudían incontrolablemente). La conclusión que la comunidad científica extrae es que los resultados son tan inciertos que se debería limitar este tipo de experimentos al laboratorio. Por otra parte, podría estudiarse y si mostraran eficacia usarse para transplante celular, las células provenientes de fetos abortados espontáneamente.
Obtención de células madre por clonación o por división partenogénica de óvulos
 Los experimentos de clonación que permitieron el nacimiento, en 1997, de la oveja "Dolly", se llevaron a cabo por transferencia del núcleo de una célula somática de la ubre de una oveja a un oocito desnucleado (Wilmut I., et al.,1997) de otra. Se demostró así que la información genética del núcleo de una célula diferenciada puede ser artificialmente reprogramada y desdiferenciada hasta recuperar la información del cigoto: célula totipotente capaz de emitir el mensaje genético completo y dar origen a un nuevo individuo. Este logro con la oveja planteó la posibilidad de generar in vitro células troncales con potencial terapéutico a partir de un número pequeño de células diferenciadas del paciente a tratar (por ejemplo una muestra de piel o biopsia muscular), sin el problema de rechazo inmune de cualquier transplante que proceda de células de donante.
Los experimentos de clonación que permitieron el nacimiento, en 1997, de la oveja "Dolly", se llevaron a cabo por transferencia del núcleo de una célula somática de la ubre de una oveja a un oocito desnucleado (Wilmut I., et al.,1997) de otra. Se demostró así que la información genética del núcleo de una célula diferenciada puede ser artificialmente reprogramada y desdiferenciada hasta recuperar la información del cigoto: célula totipotente capaz de emitir el mensaje genético completo y dar origen a un nuevo individuo. Este logro con la oveja planteó la posibilidad de generar in vitro células troncales con potencial terapéutico a partir de un número pequeño de células diferenciadas del paciente a tratar (por ejemplo una muestra de piel o biopsia muscular), sin el problema de rechazo inmune de cualquier transplante que proceda de células de donante.
Como veremos más adelante, el estado del material genético propio de cada individuo no es el mismo al inicio, que a lo largo del desarrollo, o que en cada tipo celular en los diferentes órganos y tejidos del nacido y adulto. En el inicio, tras la fecundación, el material genético recién construido del nuevo individuo tiene la “impronta” paterna y materna, propia de los gametos de sus progenitores. Los cromosomas se estructuran de diferente manera y además el DNA sufre una serie de modificaciones químicas en el proceso de desarrollo y maduración del organismo. Hay una “programación” de la información genética a lo largo de la vida. Por ello clonar un adulto, o simplemente un embrión de varias semanas, supone “reprogramar” la información genética para obtener el estado que de forma natural tiene el mensaje genético del cigoto. Es esta la dificultad técnica principal que existe actualmente. Por ello la clonación que dio origen a la oveja Dolly fue un resultado positivo entre cientos de intentos, y además no se pudo “desprogramar” por completo la información genética. Por ello está oveja era adulta (“vieja”) al nacer; tenía la edad de la que sacaron el núcleo de una células de la ubre a partir de la cual fue clonada.
Conocido con el ambiguo nombre de "clonación terapéutica" este tratamiento supone la creación de un hermano clónico del paciente, que se destruiría en una etapa inicial de su desarrollo embrionario, la de blastocisto de 5 días, para convertirse en donante de las células de su organismo; por ser un clon del paciente, con igual dotación genética que él, evitaría el problema del rechazo del injerto celular. La clonación terapéutica está encaminada a crear embriones para ser utilizados como material con fines terapéuticos.
Recientemente se ha publicado un artículo (Cibelli J.B. et al, 2001) acerca del “intento fracasado” de obtener embriones humanos clónicos. El experimento no ha logrado su objetivo: ni un solo blastocisto, embrión de 5 días que tiene ya formadas las células madre buscadas. Consiguieron 71 óvulos de 7 mujeres jóvenes, que tuvieran al menos un hijo biológico y de ellos usaron 57 que estaban maduros en el momento de la extracción.
Con la mitad de estos óvulo se indujo químicamente una transformación que convirtió a cada óvulo en una célula (que llaman embrión pronuclear) capaz de dividirse. Ninguno de los 6 “falsos embriones”, que consiguieron mantener incluso 7 días en cultivo, tenía células madre embrionarias. Era evidente que ninguno era un embrión: la partenogénesis o multiplicación, sin más reprogramación del material genético, sólo genera un puñado de células más o menos organizadas, y no un embrión. Si la partenogénesis hubiera dado el resultado buscado: inducir que el material genético doble del óvulo hubiera comenzado el desarrollo embrionario, como si dicho óvulo hubiera sido fecundado, se hubiera obtenido un blastocisto.
A los otros óvulos (unos 19) consiguieron quitarles su núcleo y transferirles el núcleo de células humanas de adulto (bien de fibroblastos o de las células que acompañan al óvulo), parecido al método que se usó para producir la oveja Dolly), aunque sin conseguir tampoco una buena reprogramación. Once no se dividieron, ni siquiera a dos células y otros seis se dividieron hasta 4 o hasta 6 células y nada más. No eran realmente “embriones tempranos” sino un grumo de células (llamamos embrión temprano en general al embrión antes de la anidación en el útero, de unas dos semanas de vida; y más en concreto se guarda ese nombre genérico de temprano al de pocos días, antes de tener ya la forma de blastocisto). Además estos falsos embriones no “sirven”: carecen de las codiciadas células madre embrionarias.
Ahora bien, si la transferencia de núcleo hubiera dado el resultado buscado: un blastocisto clónico del donante del núcleo, hubiera podido desarrollarse para dar un embrión de una, dos, cuatro... células y blastocisto, como muestran las fotografías siguientes.
Este experimento de intento de clonación ha tenido muchas críticas y rechazo no sólo porque ha fracasado sino porque para algunos no es necesario clonar embriones, ya que se dispone de los embriones producidos por fecundación in vitro, que sobran. Puestos a producir embriones, bien de “sobra” de la fecundación in vitro, bien clones del paciente, hay que reconocer que estos últimos serían mejor “material biológico”, por estar preparados “a la carta”. Y además en ambos casos habría que disponer de mujeres jóvenes donantes de óvulos. Llama la atención las campañas publicitarias (ofreciendo una compensación económica) de donantes de óvulos, como gesto solidario con mujeres infecundas que deseen tener hijos. Si es una donación solidaria, sería la propia futura madre uterina la que buscaría una amiga o familiar para ser la madre biológica de su futuro hijo. Este experimento muestra que las donantes para lo que son necesarias es para investigación.
Hoy sabemos muy bien que esos millares congelados son embriones humanos; que las células madre que obtengamos a partir de ellos no son fácilmente controlables; y que cuando sepamos orientarlas para dar neuronas, o células productoras de insulina, habrá que conseguir que no produzcan rechazo por parte del paciente. Demasiadas inseguridades, reales, para las expectativas creadas en miles de enfermos y miles de familias. Y sobre todo estas células embrionarias pluripotenciales, que pudieran diferenciarse in vitro, y que podrían suponer un potencial terapéutico, llevaría siempre consigo la destrucción de embriones humanos.
Por ello, la decisión de usar o no usar para esta investigación las decenas de miles de embriones congelados que parecen existir en diferentes países, es muy delicada. Ciertamente esos embriones tendrán que dejarse morir. Ciertamente se ha creado una expectativa de curación de enfermedades. Pero usarlos es abrir una puerta falsa para la medicina. Se trata de que estos seres humanos de pocos días no “sobren”, de no producir embriones, ni por fecundación in vitro ni por clonación, para investigar ahora y para usarlos más adelante como material terapéutico. Una vida humana debe ser respetada siempre; un ser humano no debería ser jamás un medio, para un fin por bueno que sea ese fin.
__________________________
Bibliografía (V)
Cibelli, J.B., Kiessling, A.A., Cunniff, K. et al. Somatic cell nuclear in humans: Pronuclear and early embryonic development. The Journal of Regenerative Medicine 2001; 2:25-31.
Freed, C.R., Green, P.E., Breeze, R.E. et al. Transplantation of embrionic dopamine neurons for severe Parkinson"s disease. N. Engl. J. Med. 2001; 33:710-719.
lt female canary brain. Proc. Natl. Acad. Sci. USA 1983; 80:2390-2394.
Gussoni, E., Soneoka, Y., Strickland, C.D. et al. Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 1999; 401:390-394.
Itskovitz-Eldor, J., Schulding, M., Karsenti, D. et al. Differentiation of human embryonic stem cells into embryoid bodies comprising the three embryonic germ layers. Mol. Med. 2000; 6:88-95.
Lopez-Lozano, J.J.; Mata, M., Bravo, G. Transplantes neurales en la enfermedad de Parkinson: resultados clinicos tras 10 anos de experiencia. Grupo de Trasplantes Neurales de la CPH. [Neural transplants en Parkinson disease: clinical results of 10 years of experience. Group of Neural Transplants of the CPH]. Rev. Neurol. 2000; 30:1077-1083.
Shamblott, M.J., Axelman, J., Littlefield, J.W. et al. Human embryonic germ cell derivatives express a broad range of developmentally distinct markers and proliferate extensively in vitro. Proc. Natl. Acad. Sci. USA 2001; 98:113-118.
Shamblott, M.J., Axelman, J., Wang, S. et al. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc. Natl. Acad. Sci. USA 1998; 95:13726-13731.
Studer, L., Tabar, V., McKay, R.D.G. Transplantation of expanded mesencephalic precursors leads to recovery in Parkinsonian rats. Nature Neuroscience 1998; 1:290-295.
Wilmut, I., Schnieke, A.E., McWhir, J. et al. Viable offspring derived from fetal and adult mammalian cells. Nature 1997; 385:810-813.
_____________________________________
VI.- El arranque a vivir: el tiempo cero
La fecundación: el diálogo celular y molecular de los gametos paterno y materno
El proceso que constituye un nuevo ser humano (como de cualquier otro viviente mamífero) es el proceso de fecundación. Con él se prepara la materia recibida de los progenitores para dar una unidad celular con el fenotipo (las características) propias de inicio o arranque. Esto es, con capacidad de comenzar a emitir, o expresar, el mensaje genético del nuevo individuo. En la fecundación natural, en el engendrar de los padres, el proceso acabaría en la formación de una celular con un fenotipo característico de la que se denomina cigoto.
Esta célula, embrión iniciando su vida, es más que la fusión del gameto aportado por el padre y el aportado por la madre. A lo largo de un laborioso y armónico proceso el material genético de ambos se prepara, se modifica estructural y químicamente, se funden fragmentos de diferentes tipos de membranas de ambos gametos para dar la peculiar membrana del cigoto y los diversos componentes del interior de las células se ordenan de forma adecuada para la primera división celular, con la que arranca a vivir, convirtiéndose en embrión bicelular.
El proceso de fecundación de mamíferos es bien conocido. Las primeras etapas se suceden en orden al encuentro de los gametos. En ellas ambos se van activando y terminando la maduración final que les capacita para construir el embrión cigoto.
- 1. El del espermio, gameto masculino, es atraído hacia las trompas uterinas y capacitado (es decir “limpiado” de los componentes que ocultan los receptores de reconocimiemto del óvulo) en el tracto genital femenino.
- 2. De esta forma se produce el reconocimiento específico entre el espermio, maduro y capacitado, y óvulo maduro, a través de proteínas presentes en la zona pelúcida, o cubierta que rodea al óvulo, y presentes en la membrana externa de la cabeza del espermio.
- 3. Esta interacción activa al espermio. La célula germinal masculina sufre la llamada reacción acrosómica, que le permite liberar el contenido de la gran vacuola, el acrosoma. El contenido consiste en enzimas capaces de ir abriendo un canal en la trama de la zona pelúcida del óvulo, y de esta forma avanzar por ella.
- 4. Cuando el espermio activado atraviesa la zona pelúcida interactúa con la membrana del el óvulo y le activa. La interacción entre la membrana del espermio y la membrana plasmática del óvulo produce un aumento de los iones calcio en la zona del óvulo por la que han entrado en contacto. Se denomina reacción cortical a la salida de componentes de los granos corticales del óvulo que hace rígida la zona pelúcida (reacción de la zona) e impide así que pudieran entrar nuevos espermios. La elevación local del calcio constituye la base molecular del control de las siguientes etapas: el calcio hará que se formen filamentos contráctiles en dicha zona que tiran hacia dentro del núcleo del gameto paterno. A la vez el calcio pone en marcha la síntesis de proteínas que hasta ese momento estaba detenida en el óvulo maduro y ese mismo ion calcio organiza los pronúcleos paterno y materno.
- 5. La etapa final de la fecundación es la constitución del cigoto con la entrada del núcleo del espermio en el oocito. El óvulo maduro es una célula asimétrica, con el núcleo cercano a la membrana delimitando una zona o casquete con diferente composición del resto; el espermio interacciona con la membrana del óvulo por una zona situada siempre fuera de este casquete. En esta zona se produce la elevación el calcio que difunde al resto de la célula y la organiza según un eje preciso en la estructura celular de cigoto.
El encuentro, preparación y fusión de los pronúcleos paterno y materno, colocados en la superficie, para dar el núcleo del cigoto, que se sitúa en el centro del cigoto, es un lento proceso perfectamente acompasado en el tiempo y en el espacio. El DNA de cada pronúcleo está reestructurado, y con la impronta parental, materna o paterna, (grado de metilación de citosinas ) específica y propio de células germinales.
- a) En primer lugar se termina la meiosis del oocito. Es decir elimina la mitad de su dotación genética (que era diploide en el óvulo maduro) y queda esa otra mitad formando el pronúcleo materno (haploide).
- b) A continuación se produce la descondensación y del pronúcleo paterno quedando el DNA en situación de poder expresar la información genética. Al mismo tiempo la cromatina del pronúcleo materno comienza a programarse de acuerdo con la estructura y química propia de un mensaje genético que va a empezar una nueva emisión del mensaje; esto es va perdiendo ya la impronta propia de gameto materno.
- c) El pronúcleo paterno atrae al materno y se mezclan y organizan en una unidad en el centro del cigoto. Los dos pronúcleos, son ya el núcleo, que porta el patrimonio genético del hijo. Queda por tanto el cigoto preparado para la primera división tras la duplicación del DNA.
Es interesante notar que la dotación genética paterna y materna aporta cada una de las dos mitades de los 23 pares de cromosomas que constituyen el patrimonio genético de cada ser humano. Pero ambas mitades son asimétricas en lo que se refiere a su estado de “impronta” y en cuanto a estructuración del material cromosómico. Este hecho marca además otro muy importante: el desarrollo embrionario, y después en la vida extrauterina y adulta, la contribución paterna y materna es selectiva y se requieren ambas completas para la construcción y funcionamiento del organismo. La biología pone así una barrera natural para que necesariamente cada individuo sea hijo de uno y una.
Obtención del cigoto por fecundación in vitro
El cigoto puede ser bien constituido desde el punto de vista biológico en un proceso de fecundación en el laboratorio que se limitara a “acercar” los gametos masculinos, concentrados y capacitados previamente, a un óvulo maduro en un medio de cultivo que imita las condiciones fisiológicas de las trompas uterinas. De esta forma sólo los gametos dotados genéticamente de manera correcta, podrían producir una buena fecundación. La práctica clínica es, habitualmente, mucho más agresiva.
Como se ha señalado, la fecundación eficaz, en tanto proceso constituyente de un nuevo viviente, es un proceso que exige unas condiciones sumamente precisas; una de las principales se refiere al estado de maduración del óvulo: la que conlleva un ciclo natural. Es conocido que, para aumentar la eficacia de las técnicas de fecundación asistida de humanos, se suele inducir una multiovulación; un estudio reciente (Human Reproduction. Vol. 16, pag. 221-225) muestra que los embriones humanos originados por fecundación de óvulos que proceden de una multiovulación tienen más dificultad para anidar y, los que lo consiguen se desarrollan con más malformaciones que los originados por fecundación del óvulo madurado de forma natural en un ciclo; más aún, la madre por efectos del fármaco que se usa en estos casos aporta un microentorno más agresivo al embrión que trata de anidar.
Si a esto se suma que, en la mayor parte de las clínicas de Fecundación Asistida, la fusión de los gametos se realiza por inyección directa del espermio dentro del óvulo, se comprende que la viabilidad del embrión producido sea siempre mucho menor que la del engendrado.
Las dificultad de que la fecundación “forzada” de lugar a un cigoto perfectamente polarizado, como se muestra en el esquema siguiente, es una llamada de atención a la práctica clínica: se producen embriones con que no tienen las condiciones ambientales requeridas para constituirse y desarrollarse con normalidad. Por el contrario un embrión engendrado, en su entorno natural por tanto, puede sobrevivir; de hecho se conoce desde hace tiempo que los abortos tempranos espontáneos son mayoritariamente de embriones con malformaciones, y muy raramente son embriones bien formados.
El cigoto se forma en la fecundación natural de un óvulo maduro por un espermio, que entra por una zona definida (amarillo). El cigoto está perfectamente organizado y la primera división para dar el embrión de dos células se produce según un plano fijo. Si la fecundación in vitro se hace por microinyección del espermio a un óvulo, o si estos no están maduros, el cigoto resultante no está perfectamente organizado.
Embrión clonado
En la fecundación el material genético aportado por los gametos paterno y materno se encuentra en el estado adecuado: tanto el espermio como el óvulo ha sufrido la correspondiente gametogénesis, y tanto las membranas, como los pronúcleos son capaces de dar las diferentes etapas que conducen a la constitución del cigoto.
Sin embargo, en la construcción del material celular inicial por clonación no hay una fusión de gametos sino una transferencia del material genético de una célula a otra. Este material nuclear ha de ser inducido al estado en que se encuentra en la fecundación natural el núcleo proveniente de los gametos paternos (Kubiak J.Z: and Johnson, BioEssays 23, 359-364, 2001). La célula receptora debe ser un óvulo ya que el contenido citoplásmico, capaz de aportar las una organización asimétrica y una composición molecular de la membrana debe corresponder con la del óvulo maduro. Y a su vez debe ser activado, mediante una elevación de la concentración de iones calcio en el su citoplasma; por ello se requiere someter a la célula “hibrida” a pulsos de calcio a fin de activar el inicio del desarrollo.
Como ya se ha comentado el proceso de “reprogramación” de esta célula mixta, para alcanzar las características enormemente precisas del cigoto, no es nada sencillo. La posibilidad de clonación por transferencia de núcleo requiere reprogramación (desdiferenciación o rejuvenecimiento) del código genético de la célula que aporta el nucléo.
Reprogramación que técnicamente no está aún conseguida como muestra el envejecimiento prematuro desde el nacimiento de la oveja Dolly; más aún, se ha descrito (Han, Yong- Manh, Nature Genetics, vol 28, p. 173, 2001) que la frecuencia de muertes en fase embrionaria y las anomalías que presentan los embriones obtenidos por clonación mediante transferencia de un núcleo, se debe a que el DNA, por su situación, sufre un anormal o erróneo proceso de metilación y desmetilación en las primeras etapas. Es decir, carece de la “impronta” propia, elaborada a partir de la impronta parental. El clon es un hermano del donante del núcleo. Y este núcleo de una célula de un individuo neonato o adulto, ha borrado durante su desarrollo la impronta heredada de sus progenitores en el momento de su concepción.
Dos gemelos idénticos de una misma fecundación
Habitualmente se considera que los hermanos gemelos idénticos, que proceden de la fecundación de un óvulo por un espermio, necesariamente se forman por separación física, en dos unidades celulares, de igual o diferente número de células cada una. Esta separación de algunas células para constituir dos, o incluso tres, agrupaciones permite que emitan por separado y unitariamente un mensaje genético “que dice” lo mismo pero que está escrito en cada caso sobre unos cromosomas del núcleo de células diferentes. Comentaremos después esta posibilidad. Ahora interesa hacer notar que el proceso de fecundación puede terminar en dos cigotos.
En efecto, los procesos que como la fecundación, tienen etapas que se suceden en el tiempo con una sincronización temporal y también espacial son denominados procesos “complejos” y se caracterizan por tener una dinámica “caótica”. Esto no significa imperfección, ni anomalía. Es natural: el lado irregular de la naturaleza. Significa que muy infrecuentemente, y por las fluctuaciones naturales de la difusión del calcio, el proceso de división celular se puede adelantar en el tiempo al resto de las etapas que completan la fecundación: son dos células, entonces, y no una sola las que acaban estas etapas y terminan con el fenotipo propio del cigoto.
__________________________
Bibliografía (VI)
Amit, M., Carpenter, M.K., Inokuma, M.S., Chiu, C.P., Harris, C.P., Waknitz, M.A., Itskovitz-Eldor, J., Thomsom, J.A. (2000) Clonally derived human embryonic stem cells lines maintain pluripotency and proliferative potential for prolonged periods of culture. Developmental Biology 227, 271-278.
Beddington, R.S.P., Robertson, E.J. (1999) Axis development and early asymmetry in mammals. Cell 96, 195-209.
Bedford JM (1977) Sperm-egg interaction: The specificity of human spermatozoa. The Anatomical Record 188, 477-487.
Beford JM, Cooper GW (1978) Membrane fusion events in fertilization of vertebrate eggs. En: Poste G, Nicolson GL (eds). Membrane Surface Reviews (Membrane Fusion). vol. 5, North-Holland, Amsterdam, pp. 65-125.
Capmany, G., Taylor, A., Braude, P.R., Bolton, V.N. (1996) The timing of pronuclear formation, DNA synthesis and cleavage in the human 1-cell embryo. Molecular Human Reproduction 2, 299-306.
Carlson, B.M. (1999) Human Embryology and Developmental Biology, 2nd edition. Mosby, St. Louis.
Carroll J. (2001) The initiation and regulation of Ca2+ signalling at fertilization in mammals. Semin Cell Dev Biol. 12:37-43.
Edwards, R.G., Beard, H.K. (1997) Oocyte polarity and cell determination in early mammalian embryos. Molecular Human Reproduction 3, 863-905.
Evans JP, Foster JA, McAvey BA, Gerton GL, Kopf GS, Schultz RM (2000) Effects of perturbation of cell polarity on molecular markers of sperm-egg binding sites on mouse eggs. Biology of Reproduction. 62, 76-84.
Gyllensten U, Wharton D, Josefsson A, Wilson AC (1991) Paternal inheritance of mitochondrial DNA in mice. Nature 1991 Jul 18;352(6332):255-257.
Hirao, Y., Hiraoka, J. (1987) Surface architecture of sperm tail entry into the hamster oocyte. Development, Growth and Differentiation 29, 123-132.
Howlett, S.K., Bolton V.N. (1985) Sequence and regulation of morphological and molecular events during the first cell cycle of mouse embryogenesis. Journal of Embryology and Experimental Morphology 87, 175-206.
Levron, J S Munne, S Willadsen, Z Rosenwaks and J Cohen (1995) Male and female genomes associated in a single pronucleus in human zygotes. BIOLOGY REPRODUCTION 52 653-657.
Naish, S.J., Perreault S.D., Zirkin, B.R. (1987a) DNA synthesis in the fertilizing hamster sperm nucleus: sperm template availability and egg cytoplasmic control. Biology of Reproduction 36, 245-253.
Naish, S.J., Perreault S.D., Zirkin, B.R. (1987b) DNA synthesis following microinjection of heterologous sperm and somatic cell nuclei into hamster oocytes. Gamete Research 18, 109-120.
Pedersen, R.A. (2001) Sperm and mammalian polarity. Nature 409, 473-474.
Phillips, D.M., Shalgi, R. (1980) Surface architecture of the mouse and hamster zona pellucida and oocyte. Journal of Ultrastructural Research 72, 1-12.
Piotroska, K., Zernicka-Goetz, M. (2001) Role for sperm in spatial patterning of the early mouse embryo. Nature 409, 517-521.
Sathananthan, A.H., Kola, I., Osborne, J., Trounson, A:, Bongso, A. Ratnam, S.S. (1991) Centrioles in the beginning of human development. Proceedings of the National Academy of Sciences of the USA 88, 4806-4810.
Telford, N.A., Watson, A.J., Schultz, G.A. (1990) Transition from maternal to embryonic in early mammalian development: a comparison of several species. Molecular Reproduction and Development. 26, 90-100.
Tesarik, J. y Kopecny, V. (1989a). Development of human male pronucleus: ultrastructure and timing. Gamete Research 24, 135-149.
Tesarik, J., Kopecny, V. (1989b) Nucleic acid synthesis and development of human male pronucleus. Journal of Reproduction and Fertility 86, 549-558.
Visconti PE, Kopf GS. (1998) Regulation of protein phosphorylation during sperm capacitation. Biology of Reproduction 59:1-6.
Wilcox AJ, Weinberg CR, Baird DD (1995) Timing of sexual intercourse in relation to ovulation - Effects on the probability of conception, survival of the pregnancy and sex of the baby. New England Journal of Medicine 333, 1517-1521.
Winkel, G.K. and Pedersen, R.A. (1988) Fate of the inner cell mass in mouse embryos as studied by microinjection of lineage tracers. Dev. Biol. 127, 143–156.
____________________________
VII.- Diálogo molecular madre-hijo en la primera semana de vida
Las tareas del embrión en su primera semana de vida
La construcción del organismo se inicia con una etapa de crecimiento (multiplicación del número de células por división de cada una en dos), que tiene perfectamente acompasada su velocidad con la velocidad de aparición de componentes específicos de las membranas celulares del embrión bi, tetra, octocelular, etc. El reconocimiento y la interacción específica de estos componentes mantienen las células resultantes de la multiplicación unidas en un conjunto no sólo físico sino también funcional; esto es, una unidad orgánica.
El dinamismo propio de la emisión del mensaje configura la materia en este estadio sincronizando de crecimiento y organización multicelular. La activación inicial (fundamentalmente producida por la elevación de calcio en el citoplasma del cigoto) “libera” o desbloquea la información contenida en él. A su vez, la interacción célula-célula activa las señales intracelulares modificando el estado del genoma: informan a cada de las células de su identidad como parte de un todo bicelular, tetracelular, y así sucesivamente. Una vez producida la primera división se origina el embrión bicelular y posteriormente tetracelular.
 Hasta el estadio de 8 células las blastómeras conforman un grupo de células asociadas. Sin embargo, a partir de la tercera segmentación, los blastómeros realizan al máximo sus contactos entre ellos formando una grupo compacto de células mantenido por uniones estrechas. Este proceso, conocido como compactación, separa una porción de células internas, que se comunican entre sí por uniones en hendidura, de otras células que se disponen exteriormente. Aproximadamente 3 días después de la fecundación, las células del embrión compactado se dividen otra vez para formar una mórula de 16 células. Las células internas de la morula constituyen la masa celular interna (MCI), y las células que rodean a estas constituyen la masa celular externa. Las células de la MCI dan origen a tejidos del embrión propiamente dicho, y las células externas forman el trofoblasto, que más tarde contribuye a la placenta.
Hasta el estadio de 8 células las blastómeras conforman un grupo de células asociadas. Sin embargo, a partir de la tercera segmentación, los blastómeros realizan al máximo sus contactos entre ellos formando una grupo compacto de células mantenido por uniones estrechas. Este proceso, conocido como compactación, separa una porción de células internas, que se comunican entre sí por uniones en hendidura, de otras células que se disponen exteriormente. Aproximadamente 3 días después de la fecundación, las células del embrión compactado se dividen otra vez para formar una mórula de 16 células. Las células internas de la morula constituyen la masa celular interna (MCI), y las células que rodean a estas constituyen la masa celular externa. Las células de la MCI dan origen a tejidos del embrión propiamente dicho, y las células externas forman el trofoblasto, que más tarde contribuye a la placenta.
Es significativo que la segunda división, la de cada uno de estos blastómeros no se realiza exactamente simultánea en el tiempo en cada uno de ellos. Hay un estadio de tres células y las procedentes del primer blastómero se colocaran hacia el interior de la mórula y serán las células de la masa interna del blastocisto.
A la vez que se van produciendo estas divisiones comienzan a aparecer ordenadamente glicoesfinfolipidos y glicoproteínas de membrana que mantiene el orden celular y señalan a la célula la posición que ocupa en el embrión bi, tetra, u octocelular y estableciendo los ejes del embrión. Con las primeras divisiones se forma la mórula. Se constituyen como un mosaico de células que pueden distinguirse por diversos marcadores que señalan el destino que seguirán después; así, no es asunto de azar entrar a formar parte de la zona interna o externa del embrión de 8 o de l4 células, o más tarde de la masa celular interna, o del trofoblasto del blastocisto. Además de los “pegamentos” específicos de las diferentes etapas en las membranas celulares, la asimetría ya de la primera división y la diferente velocidad de la segunda división (que dará el embrión tricelular) y de la tercera división (que dará el embrión tetracelular) les hace tener una historia diferente (espacial y temporal) y una polarización diversa puntos clave para un crecimiento orgánico. No es el embrión temprano, un tejido homogéneo e indiferenciado: pueden distinguirse por marcadores que además señalan el destino que seguirán.
Por tanto, las células resultantes de esas primeras divisiones del cigoto no son un simple amasijo de células vivas, semejantes entre sí y semejantes al cigoto y dotadas cada una de la misma individualidad que éste. A diferencia de lo que sería un grupo de células vivas encerradas bajo una cubierta esférica, sin más relación entre sí que la mera cercanía física, las células del embrión temprano constituyen una única realidad biológica, forman ya un elementalísimo organismo bicelular, tetracelular, etc. Esa unidad procede del hecho de que están comunicadas entre sí. Esas uniones que amarran a las células entre sí hacen que cada una de éstas sintetice y mantenga en su interior señales moleculares que les dan noticia a cada una de la presencia de las otras y les dicen además cómo seguir adelante. Se establece así una conexión de gran precisión entre las células que las ordena en la arquitectura propia de esa etapa.
Los “pegamentos”, algunos de ellos ausentes por completo en las células germinales de sus progenitores, aparecen en un momento preciso, y desaparecen después, también en un momento preciso. Puede decirse que el embrión en este periodo tan temprano de la vida -de unos seis días si se trata de un embrión humano- sólo tiene que ocuparse de seguir estas instrucciones. La madre acumuló en el óvulo alimentos y energía que permitirán al embrión vivir, mientras recorre el largo camino del oviducto que va del extremo superior de la trompa, donde habitualmente comienza su vida, hasta el útero, donde se implantará para seguir recibiendo ayuda hasta estar en condiciones de nacer.
Durante las primeras etapas de desarrollo (los primeros días de vida) el embrión tiene un tamaño de 0.1 - 0.15 mm. En este periodo inicial vivir es fundamentalmente crecer: no en el sentido de aumentar de tamaño, cosa que no podría hacer por estar rodeado de una especie de caparazón, la zona pelúcida, que rodeaba al óvulo del que procede, sino multiplicando el número de células por divisiones sucesivas de la primera, el cigoto. En esta proliferación celular cooperan tanto el embrión como la madre. Tras la primera división, y una vez que las moléculas de adhesión dejan bien “pegadas” las dos primeras células, éstas reciben instrucciones para elaborar otra molécula concreta, la de un receptor, que sitúan en la membrana. Este receptor reconoce, y capta, una molécula de elaboración materna, un factor de crecimiento que insta a una nueva división. Sintetizan este factor de crecimiento las células maternas, primero las de las trompas, y después las del útero, promoviendo en este caso la multiplicación de las células de la masa interna del embrión ya en fase de blastocisto anidado.
La madre provee los factores necesarios para el desarrollo del embrión, a su paso por las trompas hacia el útero. La fecundación artificial priva al embrión de varios días de las ventajas del entorno materno, disminuyendo así su capacidad de sobrevivencia.
Al finalizar la primera semana de desarrollo, el embrión “eclosiona” (se libera) de la zona pelúcida, agranda ligeramente su tamaño (por un incremento del número de células) y comienza el proceso de implantación (anidación) en la pared uterina, proceso que se completa hacia finales de la segunda semana de desarrollo.
Gemelación natural, clonación por gemelación y quimeras
De forma no natural, sino mediante la intervención experimental en el laboratorio, teóricamente podría conseguirse la separación física de las células que constituyen el embrión temprano (hasta que alcanza unas 8 células), despegándolas unas de otras. Si se colocan en el interior de una cubierta adecuada, que remede la cubierta pelúcida natural, cada célula o grupo de células podrían seguir sus divisiones y conseguirse así varias mórulas, que tras implantarlas en uno o varios úteros, adecuadamente preparados, desembocarían en el nacimiento de gemelos clónicos, es decir, réplicas exactas del pequeño embrión inicial. Obviamente un embrión de más días, más desarrollado y avanzado, no puede ser clonado, y mucho menos un recién nacido o un adulto, por la modificación del DNA de la que ya hemos hablado: a medida que avanza el desarrollo embrionario, los genes de cada individuo guardan memoria del avance de la vida. La composición química y la arquitectura de los cromosomas cambian progresivamente de forma irreversible, abriendo unas posibilidades y cerrando otras. En el mensaje inicial, que contenía las instrucciones para que se forme ese individuo, se producen durante el desarrollo, al pasar por cada fase concreta de esa vida naciente, cambios sin retorno.
A veces ocurre una gemelación natural que posiblemente sea debida a que se separan algunas células del embrión temprano y se reagrupan de nuevo, dando lugar a dos unidades de multiplicación celular, que dará lugar a dos embriones, que anidan por separado y originan dos hermanos gemelos monocigóticos. Es importante destacar que la segregación de células no origina, ni significa, partición del embrión, ni de la información genética. Ni mucho menos significa que no sea un individuo. El que los dos individuos gemelos tengan igual información genética no les hace idénticos e indiscernibles biológicamente. Cada actualización del programa configura un ser vivo diferente, e individual, individualizando los elementos materiales con que se construye el organismo. Se individualizan con la actualización y emisión separada del mensaje genético, lo que permite constituir individualidades corporales independientes y con ello diferentes identidades biológicas. No se parte el embrión en dos, sino que una parte de él, por estar en el inicio de la emisión del mensaje, constituyen una nueva unidad de emisión.
Este tipo de gemelación, debida a que es los estados anteriores al blastocisto se separan algunas de las células y reagrupan de nuevo dando lugar a dos unidades de multiplicación celular que dará lugar a dos embriones, que anidan por separado y originan dos hermanos gemelos monocigóticos, pudiera deberse a carencia o disminución de la fuerza de pegado de las proteínas especificas de interacción celular.
Se supone que también puede ocurrir gemelación por separación en dos unidades de las células que constituyen la masa interna del blastocisto. Pues bien, la expresión de la proteína ezrina en las células más externa de la masa interna muestra que, en tal caso, se habían establecido dos procesos de emisión de mensaje genético. En efecto, células de la masa interna aisladas y cultivadas se convierten en epiteliares, ya que la ezrina se relocaliza en la corteza antes de la formación de los microtúbulos y vuelven a diferenciarse a trofoblasto.
Aunque con muy poca frecuencia puede ocurrir que en los primeros pasos del desarrollo embrionario de mamíferos se produzca una fusión embrional. Se funden en uno solo dos embriones hermanos que son gemelos heterocigóticos, engendrados simultáneamente, pero cada uno procedente de la fecundación, por espermatozoides diferentes del mismo padre, de óvulos diferentes de la misma madre. La muerte de uno de los embriones se produce cuando sus células son incorporadas por el otro en un proceso que en realidad constituye un transplante peculiar, es una quimera celular. Esto hará que el embrión receptor manifieste más adelante los caracteres propios de su hermano en las regiones de su cuerpo derivadas de las células incorporadas; algo similar a lo que ocurre en un transplante de riñón, en el que este órgano sigue manifestando los caracteres inmunológicos del donante. Ciertamente el individuo resultante de la fusión embrionaria es un híbrido, en el sentido de que los tejidos de su cuerpo serán un mosaico de células procedentes de las inicialmente suyas y de otras procedentes de las células de su hermano. Se trata de una alteración imprevisible, no determinada en el mensaje genético, del proceso normal del desarrollo en la etapa embrionaria que está posibilitada por la existencia de las moléculas de adhesión situadas en las membranas celulares que unen entre sí células de embriones de la misma edad. En este sentido un organismo mosaico es el resultado de una malformación ocurrida por accidente, sin graves consecuencias para su normal desarrollo ya que el transplante ocurre antes de que haya comenzado el proceso embrionario en que se educa al sistema inmunitario a distinguir lo propio y lo extraño y rechazar lo último.
Es posible también que este fenómeno sea debido al simple paso de un embrión a otro de una célula madre embrionaria en un estadía muy precoz del desarrollo.
En cualquier caso el proceso de gemelación o de fusión sólo se ha analizado in vitro. Los posibles mecanismos por los que se dan estos fenómenos son hipotéticos.
Otra tarea del embrión temprano: establecer con una primera diferenciación celular el tejido extraembrionario
El genoma del embrión se activa ya en el cigoto, comienza la expresión de sus genes, sincronizando el crecimiento del todo orgánico con la emisión diferencial del mensaje. Esto es, las células polares situadas en el exterior de la mórula se configuran como tejido extraembrionario: la cubierta que le permitirá el intercambio de materia, energía y señales moleculares para el crecimiento armónico con el exterior y además como primera barrera de defensa en la vida en simbiosis que iniciara con la anidación.
Así con la llegada a “blastocisto” aparecen ya dos tejidos diferenciados: el trofoblasto epidermico que funcionará como “piel embrionaria” y la masa interna que dará lugar posteriormente a los tres tejidos embrionarios. El trofoblasto no es sólo un tejido "extraembrionario", que dará lugar a la placenta, necesaria e imprescindible para la comunicación con la madre en la gestación. Es un componente del sistema inmunitario innato con un papel esencial en la defensa frente a infecciones bacterianas durante la vida intrauterina. Como trataremos en la nota siguiente, para organizar la respuesta immune en la interfase útero-placenta tiene lugar un "dialogo molecular" materno-filiar en el que el factor CSF-1 (liberado por células del sistema inmunitario de la madre presentes en las trompas) se une a receptores específicos del trofoblasto del embrión y activan dichas células.
Pues bien, para alcanzar esta primera diferenciación celular, el cigoto debe utilizar productos génicos embrionarios muy pronto en el desarrollo. A pesar de que el embrión humano la transcripción del genoma embrionario parece más marcada a partir del estadio de 8 células, existe evidencia que indica que se produce síntesis de ARN a partir del estadio cigoto (Tesarik y Kopecny, 1989; Daniels et al., 1997; Edwards y Beard, 1997). En el estadio de 8 células algunos blastomeros tienen niveles elevados de síntesis de ARN mientras otros blastomeros muestran aún el patrón mostrado por blastómeros de embriones de 4 células. Estos resultados sugieren que, además del momento en que se produce la transición del control materno al embrionario del desarrollo, parecen existir diferencias entre los blastómeros en el momento que se produce dicha transición.
___________________________
Bibliografía (VII)
Ao A, Erikson RP, Winston RM, Handyside AH. Transcription of paternal Y-linked genes in the human zygote as early as the pronucleate stage. Zygote 1994;2:281-287 .
Beddington, R.S.P., Robertson, E.J. (1999) Axis development and early asymmetry in mammals. Cell 96, 195-209.
Braude, P. Bolton V, Moore S (1988) Human gene expression first occurs between the four- and eight-cell stages of preimplantation development. Nature 332, 459–461.
Brenner, S., Dave, W., Hewrskowitz, y Thomas, R. (1990) Genes and development: molecular and logical themes. Genetics 126, 479?486.
Bursdall C.A., Damsky C.H., Pedersen R.A: (1993) The role of E-cadherin and integrins in mesoderm differentiation and migration at the mammalian primitive streak. Development 118, 829-844.
Dard N., Louvet S., Santa-María A Aghion J.,. Martin M. ,Mangeat P. and Maro B. In vivo funtional analysis of ezrin durin mouse blastocyst formación Developmental Biology 233 161-173, 2001.
Daniels, R., Lowell, S., Bolton, V., Monk, M. (1997) Transcription of tissue-specific genes in human preimplantation embryos. Human Reproduction 12, 2251-2256.
Fiddler, M., Abdel-Rahman, B., Rappolee, D.A., Pergament, E. (1995) Expression of SRY transcripts in preimplantation human embryos. American Journal of Medical Genetics 55, 80-84.
Gardner, R.L. (1968) Mouse chimaeras obtained by the injection of cells into the blastocyst. Nature 220, 596-597. Gardner, R.L. (1982) Investigation of cell lineage and differentiation in the extraembryonic endoderm of the mouse embryo. J. Embryol. Exp. Morphol. 68, 175–198. Gardner, R.L. (1997) The early blastocyst is bilaterally symmetrical and its axis of symmetry is aligned with the animal-vegetal axis of the zygote in the mouse. Development 124, 289-301. Gardner RL. The initial phase of embryonic patterning in mammals. En: Etkin LD, Jeon KW. Cell lineage specification and patterning of the embryo. Int Rev Cytol 2001;203:233-290; http://www.devbio.com/chap11/link1103.shtml; /link1104.shtml).
Graham, C.F., Kelly S.J. (1977) Interactions between embryonic cells during early development in the mouse. En: Cell Interactions in Differentiation (M. Karkinen-Jaaskelainen, L. Saxen, L. Weiss, edds). Academic Press, New York, pp. 45-57
Howlett, S.K., Bolton V.N. (1985) Sequence and regulation of morphological and molecular events during the first cell cycle of mouse embryogénesis. Journal of Embryology and Experimental Morphology 87, 175-206.
Memili, E., Dominko, T., First, N.L. (1998) Onset of transcription in bovine oocytes and preimplantation embryos. Molecular Reproduction and Development 51, 36-41.
Piko L, Clegg KB (1982) Quantitative changes in total RNA, total poly(A), and ribosomes in early mouse embryo. Developmental Biology 89, 362-378.
Piotroska, K., Zernicka-Goetz, M. (2001) Role for sperm in spatial patterning of the early mouse embryo. Nature 409, 517-521.
Rosner M.H. et al. (1991) Oct-3 and the beginning of mammalian development. Science 253, 244-245.
Sutherland, A.E., Speed, T.P., Calarco, P.G. (1990) Inner cell allocation in the mouse morula: The role of oriented division during fourth cleavage. Developmental Biology 137, 13-25.
Tarkowski A.K., Wroblewska, J. (1967) Development of blastomeres of mouse eggs isolated at the 4- and 8-cell stage. Journal of Embryology and Experimental Morphology 18, 155-180.
Telford, N.A., Watson, A.J., Schultz, G.A. (1990) Transition from maternal to embryonic in early mammalian development: a comparison of several species. Molecular Reproduction and Development. 26, 90-100.
Wiley L.M. (1984) Cavitation in the mouse preimplantation embryo: Na/K ATPase and the origin of nasceent blastocoel fluid. Developmental Biology 105, 330-342.
_____________________________________________
VIII.- Preparando el reconocimiento del padre: segunda semana de vida.
La anidación en el útero materno
En los cuatro o cinco primeros días de vida, mientras el embrión, blastocisto, se mueve a lo largo del oviducto hacia el útero, se expande dentro de la zona pelúcida. Durante esta etapa, la zona pelúcida evita que el blastocisto se adhiera a la pared del oviducto. Cuando el embrión llega al útero, debe “eclosionar” de la zona de modo de poder adherirse a la pared uterina. La implantación del blastocisto humano comienza hacia finales de la primera semana y se completa hacia el final de la segunda semana. Aproximadamente 6 días después de la fecundación (día 20 de un ciclo menstrual de 28 días), el blasocisto se adhiere al epitelio endometrial, generalmente a través del polo embrionario, el polo que contiene la masa celular interna.
Tan pronto como se adhiere al epitelio endometrial, el trofoblasto comienza a proliferar rápidamente y se diferencia gradualmente en dos capas. A continuación, y también aproximadamente en el día 6, unas extensiones del trofoblasto se extienden a través del epitelio del endometrio e invaden el tejido materno. Hacia finales de la primera semana, el blastocisto está implantado superficialmente en la capa compacta de endometrio y obtiene su nutrición de los tejidos maternos erosionados: las células del estroma materno que se encuentran alrededor del sitio de la implantación se cargan de glucógeno y de lípidos.
El embrión comienza a producir una hormona (la gonadotrofina coriónica humana, hCG) que entra a la circulación materna. Esta hormona mantiene la actividad del ovario durante la gestación y constituye la base de los tests de embarazo.
El embrión de 10 días está totalmente embebido en el endometrio. El final de la segunda semana (días 13 y 14) se caracteriza por la aparición de las vellosidades coriónicas primarias, la primera etapa en el desarrollo de la placenta.
Terminada la anidación las células de la masa interna se han organizado como disco embrionario bilaminar. La siguiente etapa, conocida como gastrulación transforma con una segunda diferenciación celular el disco embrionario en trilaminar. A partir de las células de estas tres capas embrionarias se formaran todos los órganos y tejidos.
En la primera semana el embrión se constituye en blastocisto y comienza a anidar en el útero, y en la segunda semana mientras sigue desarrollándose hasta gástrula se embebe por completo en el endometrio materno.
Simbiosis con la madre: ni parte de ella, ni injerto extraño
La relación, o dialogo molecular, madre-hijo tiene un carácter de simbiosis. Efectivamente, diversos datos acerca de la tolerancia fetomaternal han mostrado que el embrión al implantarse establece una simbiosis. Ni se comporta como injerto, ni es una parte del cuerpo materno.
 Puesto que todos los seres humanos difieren entre sí y llevan “etiquetas” que los individualizan, los órganos transplantados de una persona a otra son rechazados, a no ser que se amordace el sistema de reconocimiento de lo propio, el sistema inmunitario. El cigoto es la célula originaria en la cual la mitad de los “marcadores de lo propio” provienen del padre, y la otra mitad de la madre. El huevo fecundado y la placenta son “mitad” extraños al organismo materno.
Puesto que todos los seres humanos difieren entre sí y llevan “etiquetas” que los individualizan, los órganos transplantados de una persona a otra son rechazados, a no ser que se amordace el sistema de reconocimiento de lo propio, el sistema inmunitario. El cigoto es la célula originaria en la cual la mitad de los “marcadores de lo propio” provienen del padre, y la otra mitad de la madre. El huevo fecundado y la placenta son “mitad” extraños al organismo materno.
En ciertos casos de procreación médicamente asistida, el embrión procede de la fecundación de un ovocito y de un espermio proveniente de donantes: él es totalmente extraño a la madre que lo lleva y puede ser rechazado.
Además, mientras que sin un tratamiento inmunosupresivo apropiado, los transplantes de órgano entre donantes y receptores incompatibles acaban en un rechazo cada vez más fuerte, de una gestación a otra, las placentas son cada vez mejor toleradas y (cada vez de mayor tamaño); los embarazos sucesivos favorecen una tolerancia inmunitaria de la madre cada vez mayor hacia los tejidos paternos. Más aún, rechazan todo injerto de órganos, incluidos aquellos procedentes de tejidos de su propio feto.
Hoy se sabe que toda una red de sustancias inhibitorias actúan localmente para mantener la tolerancia inmunológica de la madre para el niño que gesta. Para un inmunólogo “clásico”, el feto no es rechazado porque no existen marcadores de lo propio (antígenos de histocompatibilidad, HLA) en la superficie de las células que separan la madre del feto, es decir, en la superficie del trofoblasto, las células de la capa más externa de la placenta. Aunque esta hipótesis no es correcta, sin embargo, la teoría de una barrera neutra entre la madre y el feto es parcialmente verdadera.
En efecto, existe en la placenta una zona en contacto con la sangre materna en la que el trofoblasto, que forma la primera línea de separación de las vellosidades de la placenta y de la sangre materna, que no expresa ningún antígeno de histocompatibilidad, pero eso no es todo. La placenta segrega factores inmunosupresores específicos, entre ellos una proteína llamada HLA G soluble, que es capaz de inhibir los linfocitos T incluso después de la activación de sus receptores, provocando hasta su eliminación.
Presentación de los antígenos del padre
Este marcador HLA G, en la superficie del trofoblasto, presenta al sistema inmunitario de la madre, lo que en el embrión es del padre; y de esta forma el embrión es tolerado por la madre, a pesar de que tiene antígenos que pertenecen a su padre, gracias a toda una red de sustancias que inhiben localmente el sistema inmunitario, y que se sintetizan tras la presentación.
Los linfocitos B que producen anticuerpos contra estos antígenos “extraños”. Pero las moléculas liberadas por la placenta inhiben estos anticuerpos potencialmente peligrosos.
De igual forma, los linfocitos T citotóxicos y las células asesinas naturales deberían ser activadas por la presencia de moléculas heredadas del padre. De nuevo, factores inmunosupresores segregados por la placenta y por el útero inactivan las células que podrían amenazar al feto.
Por tanto, en buena medida es la HLA-G “la” molécula del mantenimiento del embarazo permitiendo al embrión vivir en una autentica simbiosis con la madre. Pero no es la única. El troflobasto humano expresa también HLA-C. Y existe un autentica “red local de citoquinas”. De hecho, el embrión y la placenta se encuentran inmersas en una multidud de citoquinas segregadas por la madre, o por el trofoblasto (la “piel” del embrión).
En las primeras etapas de vida, el embrión comienza a ayudar a la madre a tolerar lo que por pertenecer al padre lleva de diferente: un dialogo programado y muy elaborado, que “plantea cuestiones y respuestas moleculares”, se instaura entre la madre y el embrión, que prologa otros diálogos que se establecen desde el principio entre la madre y su hijo. La aceptación para gestar el hijo pasa por este dialogo “tolerante” desde el momento de las primeras fases de la vida.
_________________________________
Bibliografía (VIII):
Chenguang Xu, et al “A critical Role for Murine Complement regulator Crry in fetomaternal tolerance “ Science vol. 287 , 2000, pag. 498-501.
Guleria I y Pollard, J.D. “ The trophoblast is a component of the immune system during pregnancy” Nature Medicine 6, 2000, 589-593.
Le-Bouteiller, P “The funcionality of HLA-G is emergging” Immunol Rev. 1999, 167233-44.
 [Reproducimos un importante artículo de Robert Spaemann que trata de lo que queremos decir cuando llamamos "personas" a los hombres y a las mujeres. Trata por tanto sobre la distinción entre "algo" y "alguien". Algo que es cosa no puede ser más que cosa; y quien es sujeto de derechos no puede haber sido antes una cosa sin derechos. "No existe un tránsito paulatino desde el 'algo' a 'alguien'. Si el ser persona fuera un estado, podría surgir poco a poco. Pero si la persona es alguien que pasa por diferentes estados, entonces los supone todos.” Y en todos los estados -como el embrión de pocos días, o como el infante recién nacido, o como el anciano decrépito y con déficit neurológico- tiene la misma dignidad inviolable: "no hay ningún estado del hombre que pudiera justificar negarle su existencia terrena." Pero no todos piensan así: algunos autores han negado y niegan que todos los hombres sean personas y las consecuencias prácticas son terribles: la banalización del aborto y de la eutanasia; la penosa manipulación grosera de embriones humanos como si se tratara de guisantes... En este denso y profundo artículo, el autor se centra de modo particular en la eutanasia. Hay que argumentar y hay que hacerlo bien, a fondo, hasta el tuétano de la verdad. Publicado en Biblioteca Almudi]
[Reproducimos un importante artículo de Robert Spaemann que trata de lo que queremos decir cuando llamamos "personas" a los hombres y a las mujeres. Trata por tanto sobre la distinción entre "algo" y "alguien". Algo que es cosa no puede ser más que cosa; y quien es sujeto de derechos no puede haber sido antes una cosa sin derechos. "No existe un tránsito paulatino desde el 'algo' a 'alguien'. Si el ser persona fuera un estado, podría surgir poco a poco. Pero si la persona es alguien que pasa por diferentes estados, entonces los supone todos.” Y en todos los estados -como el embrión de pocos días, o como el infante recién nacido, o como el anciano decrépito y con déficit neurológico- tiene la misma dignidad inviolable: "no hay ningún estado del hombre que pudiera justificar negarle su existencia terrena." Pero no todos piensan así: algunos autores han negado y niegan que todos los hombres sean personas y las consecuencias prácticas son terribles: la banalización del aborto y de la eutanasia; la penosa manipulación grosera de embriones humanos como si se tratara de guisantes... En este denso y profundo artículo, el autor se centra de modo particular en la eutanasia. Hay que argumentar y hay que hacerlo bien, a fondo, hasta el tuétano de la verdad. Publicado en Biblioteca Almudi]